Phalaenopsis cochlearis
|
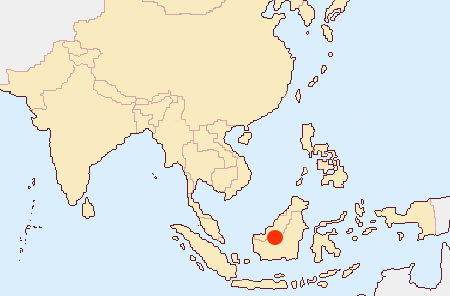
1.生息分布
| ||||
|
2.生息環境 海抜500-700m(1,500-2,000mの説がある)。気温夜間15-20C、昼間25-28C。湿度80%以上。着生あるいは岩生。多くは石灰岩にコケと共に生息していると言われることから雲霧林とおもわれる。自然での開花は9-10月。温室では初夏が多い。今日自然界ではほとんど見ることができない希少種である。 | ||
|
3.形状 3-1 花
1. 花被片 花名はリップ形状のスプーンの凹面のようなという意味。花の直径は 4cm。花被片は全体が薄緑色から写真のような黄味の強いものがあり、花弁およびラテラルセパルの蕊柱付け根に茶の斑点が入る。ラテラルセパルに斑点がないものもある。この斑点はドーサルセパルには見られないか、僅かに小さな斑点となる。 花被片は長楕円形で全体として星型である。個体差と思われるが、それぞれの花被片の先端が後ろにやや反るもの、ラテラルセパルの先端が右写真のように卵形で尖っているものなどがある。ドーサルセパルにも斑点が入るものが稀に見られる。大株になると4‐5本の花茎が発生し、それぞれが2‐3本枝分かれし、それらに2‐3輪の花をつける。 これまでに葉の様態がPhal. fuscataやPhal. kunstleriと似ており、これらのラベルで誤ってマレーシヤより入荷したことがある。香りはない。
2. リップおよびカルス 中央弁はスプーンのように凹面であり、下写真左に見られるようにクリーム色のベースに8本の茶色のストライプが入る。このストライプの本数に個体差(数の多少)は見られない。カルスは2組構成でいずれも先端が2分岐している。一方、リップ形状が比較的類似するPhal. fuscataやPhal. kunstleriの中央弁ストライプは6‐4本までであり、それぞれのストライプが太い。
3-2 さく果 Phal. fuscataやPhal. kunstleri、Phal. giganteaと子房の形状は類似する。子房の表面はPhal. cornu-cerviのさく果と比べ、艶がなく柔らかな質感があり、6本の縦筋は明瞭である。交配後、花被片は枯れ縮む。
3-3 変種および地域変異 変種は知られていないが、3-1項の写真が示すような花被片が緑色また黄色味の強いカラーフォームが稀に見られる。3-4 葉 葉は下垂性で、栽培株では一般的に葉長20cm、幅6.0 - 6.5cmの、Phal. bellinaと似た形状と厚みを持った下写真左に見られる楕円形状。一方、野生株では長楕円形や写真右のような35 - 38㎝になる葉も見られる。しばしばマーケットでは葉形状の似たPhal. kunstleriと誤って取り扱われることがある。
3-5 花茎 通常サイズの花茎の長さは20 - 30cm長で、1-2本発生し先端部に2-3輪の花を着ける。花が終了しても花茎は枯れないで翌年にその花茎の先端に花芽を着けることがあるが、枯れことも多い。下写真左は葉長35㎝以上の大株(左)と一般種2株(中央と右)を並べている。いずれもBS株であり、生育環境に依っては株サイズが大きく変化することが分かる。大株になると4-5本の花茎と、さらにそれらから分岐した茎に2-3輪が開花する。右写真は葉長の倍(50㎝)近くに伸長した、これまでに見た最長の花茎である。
3-6 根 コルク、へご板あるいはバスケットに取り付けるのが一般的である。根の伸長は緩やかである。 | ||||||||||||||||||||














|
4.育成
| |||||||||||||||||||

|
5.特記事項 野生種の入手が最も難しい原種の一つである。ボルネオ島Sarawakでの既知の生息域では絶滅しているとの2014年現地ラン園の話である。一方、フラスコ苗がタイ、台湾から販売されているが2010年いずれも純正種ではなかった経験をもつ。 自家交配に難発芽性があるのかも知れない。2011年から2014年までの間で野生種は2株しか入手できない状態となっている。野生種を交配親とする実生は現在マレーシアで入手可能である。 | ||